

Ni el mismo Darwin tenía claro cómo, a partir de la competencia por la reproducción entre los individuos, podrían surgir comportamientos altruistas, en los que uno mismo se perjudica por favorecer la reproducción de otros. Incluso hoy día existe controversia sobre la propuesta de Hamilton (1963) y Dawkins (1975) que suponen que son los genes los que compiten por multiplicarse, y que los seres vivos somos meras máquinas de perpetuación de estos genes, porque cada vez se encuentran más excepciones a esta regla. Otras teorías, como la de selección multinivel, aunque funcionalmente equivalente a la anterior, pecan de falta de planteamientos teóricos.
Nosotros proponemos una síntesis entre ellas, muy simple:
De la conocida regla de Hamilton nos quedamos con que, en todo ser vivo, existe una parte egoísta y otra altruista (Direct e Indirect fitness lo llaman, ver Fig.1). Y de la teoría multinivel entresacamos que existen presiones selectivas distintas a varios niveles de la jerarquía biológica, una que afecta a la competencia entre los individuos y otra entre grupos de individuos, y que son simultáneas, de signo contrario y de distinta intensidad.
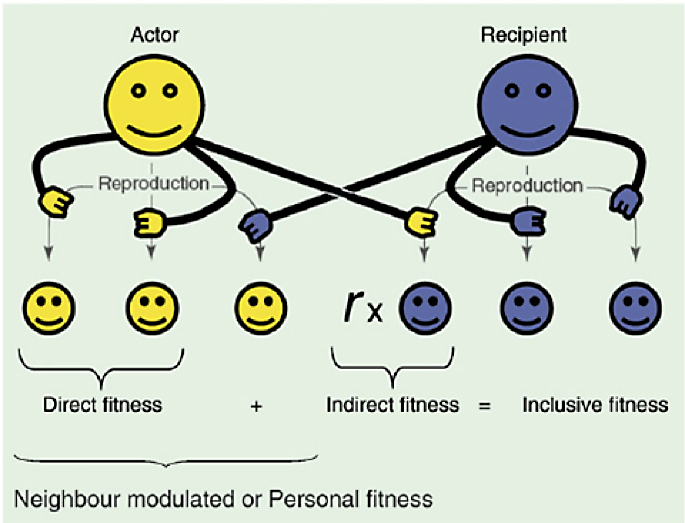
Fig. 1: La eficacia biológica inclusiva o «inclusive fitness» es la suma de su Direct y su Indirect fitness. Según demostró Hamilton, para que los genes altruistas aumenten de frecuencia, debe de existir una relación de parentesco “r” entre actor y recipiente. Figura obtenida de https://www.researchgate.net/figure/Inclusive-fitness-is-the-sum-of-direct-and-indirect-fitness-Hamilton-1964-Social_251645045
Partiendo de ello, suponemos que cualquier individuo debe de concentrar su eficacia biológica allí donde la presión selectiva es más intensa y es más probable morir. Si la presión selectiva es mayor a nivel de grupo, la vida del individuo dependerá, en mayor medida, de la supervivencia del grupo, y se seleccionarán positivamente los grupos más aptos, que estarán integrados, precisamente, por los sujetos más altruistas. Por el contrario, si la presión selectiva es mayor a nivel de individuo, se seleccionarán los sujetos que sean más egoístas.
Por ejemplo, el tigre ‒Panthera tigris‒, al no tener competencia con otros grupos de predadores, desarrollan adaptaciones individualistas, como es un gran dimorfismo sexual en tamaño corporal, para competir contra otros machos. Por el contrario, si la posibilidad de no superar la presión selectiva es más alta a nivel de grupo, solo pasarán el corte de la selección natural los grupos más aptos y los individuos más altruistas, como sucede, por ejemplo, con los animales eusociales, en los que solo prosperan adaptaciones altruistas, en beneficio del grupo, como la esterilidad o dar la vida por el grupo, o en los animales simbiontes o mutualistas obligados, que se adaptan potenciando funciones que benefician la relación simbiótica, en detrimento de las funciones vitales indispensables para su propia supervivencia.
Incluso un mismo individuo puede interactuar en un ambiente en el que exista mayor presión de grupo (o como lo llamaba Thompson (2005) “hot spots”) donde los más altruistas salen favorecidos, o puede interactuar en un “cold spots”, donde serán los egoístas los que tengan más ventajas en la reproducción. Por ejemplo, el cucaburra común ‒Dacelo novaeguineae‒ es un ave australiana que a veces se reproduce cooperativamente, dependiendo del número de nidos disponibles. Los pájaros jóvenes de una nidada anterior que no disponen de nido libre, en ocasiones ayudan a sus padres o, incluso, a otras aves mayores no emparentadas, y sacan adelante una nidada ajena. Otras cucaburras más egoístas no cooperan nunca. Pues bien, se sabe que, cuando no hay nidos disponibles, el número de pollos de las aves cooperadoras es de media mayor que las egoístas, porque en futuras nidadas se ven favorecidas con más facilidad para encontrar pareja. Como el factor ambiental que controla el nivel de presión selectiva grupal/individual es el número de nidos disponibles, a menor número de nidos disponibles, más presión de grupo, y el grupo de las aves más cooperadoras serán las más favorecidas por la selección natural. Por el contrario, a más cantidad de nidos disponibles, las aves más egoístas son seleccionadas positivamente con más probabilidad.
Si aceptamos esto, y llamamos “e” a la proporción egoísta de un individuo y “1-e” a la altruista (donde 0<e<1) y llamamos “p” a la proporción de presión selectiva entre individuos, y “1-p” a la de grupo (donde 0<p<1) podemos obtener lo que hemos dado en llamar la eficacia biológica ponderada F’:
F’ sería la aportación relativa que cada individuo hace, en términos de eficacia biológica, a sí mismo, “e”, y a su grupo, “1-e”, multiplicadas por la presión de selección relativa en cada uno de esos niveles, “p” y “1-p”, respectivamente. Así, el individuo que tenga una F’ mayor contribuirá más al nivel jerárquico que su ambiente exige y será seleccionado positivamente con más probabilidad.
La función F’ es un paraboloide hiperbólico (Fig. 2) una superficie tridimensional curva similar a una silla de montar, doblemente reglada porque se construye a partir de rectas. Tiene un punto, S, denominado “punto de silla”, donde coinciden el valor mínimo de los máximos. Si nos imaginamos este paraboloide hiperbólico como una ola, solo serán seleccionados positivamente los individuos que estén en la cresta de la ola, los de mayor F’, aunque éstos puedan tener menos Inclusive fitness (Fig. 1).
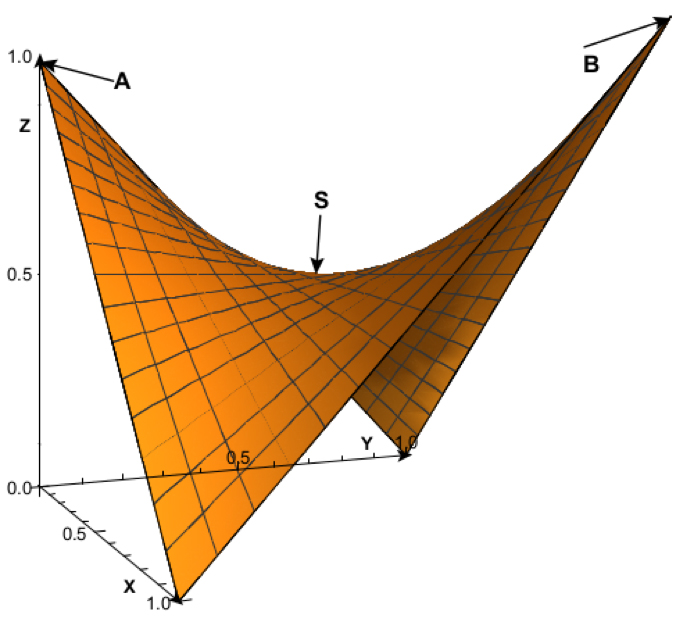
Fig. 2: Paraboloide hiperbólico de la función F’ de la eficacia biológica ponderada. El eje X es la proporción de presión selectiva individual p, el eje Y es la proporción de eficacia biológica individual e, y el eje Z corresponde al valor de F’. El punto S (0.5, 0.5, 0.5) es el punto de silla o minimax. La parábola que va del punto A (0,0,1) hasta el punto B (1,1,1) y cuyo vértice coincide con el punto de silla S, representa los valores máximos de e para cada valor dado de p. Es decir, la parábola ASB es la cresta de la ola y representa las proporciones de eficacia que serán seleccionadas positivamente para cada nivel de presión selectiva.
De este modelo destacamos que:
- Para explicar la emergencia del altruismo evolutivo no es necesario el parentesco. Simplemente con la concurrencia de individuos en un ambiente en el que la intensidad de la presión de grupo es superior a la individual, se deben de seleccionar positivamente rasgos que aumenten el altruismo. En un ambiente en el que “p < 0,5”, los individuos altruistas “e < 0,5” que trabajan para el grupo deben de dotarse de características cada vez más altruistas, porque los rasgos egoístas se convierten en deletéreos para el grupo y, por tanto, harán que los individuos tengan una F’ menor.
- Aunque parezca paradójico, solo será beneficiosa para el individuo la colaboración de sus compañeros si la presión selectiva favorece el egoísmo “p > 0,5”.
- A largo plazo, cuando predomine la presión de grupo “p < 0,5” y se mantenga este predominio en el tiempo, cada vez serán seleccionados los más altruistas “e < 0,5” llegándose al punto máximo de la cresta de la ola, el punto A (Fig. 1) donde el egoísmo es nulo y solo se vive para el grupo. Con ello se llega a la “individualización” del grupo, a la “simbiogénesis” o especiación jerárquica, o creación de una nueva especie jerárquicamente superior. Por el contrario, cuando predomina y persiste en el tiempo la presión selectiva individual, se tenderá al punto B (Fig. 1) de máximo egoísmo.
AUTOR
Javier Falgueras Cano
Premio “Accesit” en el XV Certamen Universitario Arquímedes por el trabajo «Altruismo» Realizado por Javier Falgueras Cano de la Universidad de Burgos y tutorizado por José Miguel Carretero Día.
+INFO CERTAMEN ARQUÍMEDES



No comments yet.