

Ni el mateix Darwin tenia clar com, a partir de la competència per la reproducció entre els individus, podrien sorgir comportaments altruistes, en els quals un mateix es perjudica per afavorir la reproducció d’altres. Fins i tot hui en dia hi ha controvèrsia sobre la proposta de Hamilton (1963) i Dawkins (1975), que suposen que són els gens els que competeixen per multiplicar-se, i que els éssers vius som meres màquines de perpetuació d’aquests gens, perquè cada vegada es troben més excepcions a aquesta regla. Altres teories, com la de selecció multinivell, encara que funcionalment equivalent a l’anterior, pequen de falta de plantejaments teòrics.
Nosaltres proposem una síntesi entre elles, molt simple:
De la coneguda regla de Hamilton ens quedem que, en tot ésser viu, hi ha una part egoista i una altra d’altruista (Direct i Indirect fitness en diuen, vegeu figura 1). I de la teoria multinivell traiem la conclusió que hi ha pressions selectives diferents de diversos nivells de la jerarquia biològica, una que afecta la competència entre els individus i una altra entre grups d’individus, i que són simultànies, de signe contrari i de distinta intensitat.
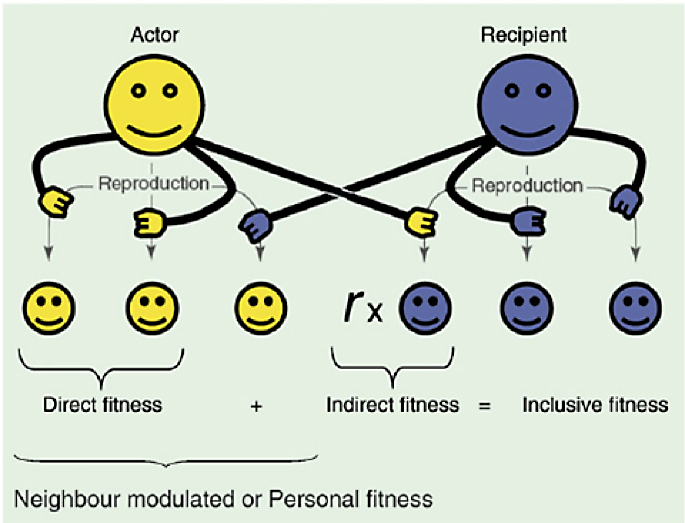
Fig. 1: L’eficàcia biològica inclusiva o «inclusive fitness» és la suma del seu «direct» i la seva «indirect fitness». Segons va demostrar Hamilton, perquè els gens altruistes augmentin de freqüència, deu existir una relació de parentiu “r” entre actor i recipient. Figura obtinguda de https://www.researchgate.net/figure/Inclusive-fitness-is-the-sum-of-direct-and-indirect-fitness-Hamilton-1964-Social_251645045
Partint d’això, suposem que qualsevol individu ha de concentrar la seua eficàcia biològica allí on la pressió selectiva és més intensa i és més probable morir. Si la pressió selectiva és més gran a nivell de grup, la vida de l’individu dependrà, en major grau, de la supervivència del grup, i se seleccionaran positivament els grups més aptes, que estaran integrats, precisament, pels subjectes més altruistes. Al contrari, si la pressió selectiva és més gran a nivell d’individu, se seleccionaran els subjectes que siguen més egoistes.
Per exemple, el tigre ‒Panthera tigris‒, com que no té competència amb altres grups de predadors, desenvolupen adaptacions individualistes, com és un gran dimorfisme sexual en grandària corporal, per a competir contra altres mascles. Al contrari, si la possibilitat de no superar la pressió selectiva és més gran a nivell de grup, només passaran el tall de la selecció natural els grups més aptes i els individus més altruistes, com ocorre, per exemple, amb els animals eusocials, en els quals només prosperen adaptacions altruistes, en benefici del grup, com l’esterilitat o donar la vida pel grup, o en els animals simbionts o mutualistes obligats, que s’adapten potenciant funcions que beneficien la relació simbiòtica, en detriment de les funcions vitals indispensables per a la seua pròpia supervivència.
Fins i tot un mateix individu pot interactuar en un ambient en què hi haja més pressió de grup (o com en deia Thompson (2005) hot espots), on els més altruistes resulten afavorits, o pot interactuar en un cold espots, on seran els egoistes els que tinguen més avantatges en la reproducció. Per exemple, el cucaburra comú ‒Dacelo novaeguineae‒ és una au australiana que a vegades es reprodueix cooperativament, depenent del nombre de nius disponibles. Els ocells joves d’una niuada anterior que no disposen de niu lliure a vegades ajuden els seus pares, o fins i tot altres aus majors no emparentades, i trauen avant una niuada aliena. Altres cucaburres més egoistes no cooperen mai. Doncs bé, se sap que, quan no hi ha nius disponibles, el nombre de pollets de les aus cooperadores és de mitjana més gran que les egoistes, perquè en futures niuades es veuen afavorides amb més facilitat per a trobar parella. Com que el factor ambiental que controla el nivell de pressió selectiva grupal/individual és el nombre de nius disponibles, a menor nombre de nius disponibles, més pressió de grup, i el grup de les aus més cooperadores seran les més afavorides per la selecció natural. Al contrari, a més quantitat de nius disponibles, les aus més egoistes són seleccionades positivament amb més probabilitat.
Si acceptem açò, i anomenem “e” la proporció egoista d’un individu i “1-e” l’altruista (on 0<e<1), i anomenem “p” la proporció de pressió selectiva entre individus i “1-p” la de grup (on 0<p<1), podem obtenir el que s’ha anomenat l’eficàcia biològica ponderada F’:
F’ seria l’aportació relativa que cada individu fa, en termes d’eficàcia biològica, a si mateix, “e”, i al seu grup, “1-e”, multiplicades per la pressió de selecció relativa en cada un d’aquests nivells, “p” i “1-p”, respectivament. Així, l’individu que tinga una F’ més gran contribuirà més al nivell jeràrquic que el seu ambient exigeix i serà seleccionat positivament amb més probabilitat.
La funció F’ és un paraboloide hiperbòlic (figura 2) una superfície tridimensional corba similar a una sella de muntar, doblement reglada perquè es construeix a partir de rectes. Té un punt, S, denominat punt de sella, on coincideixen el valor mínim dels màxims. Si ens imaginem aquest paraboloide hiperbòlic com una onada, només seran seleccionats positivament els individus que estiguen a la cresta de l’onada, els de més F’, encara que aquests puguen tenir menys Inclusive fitness (figura 1).
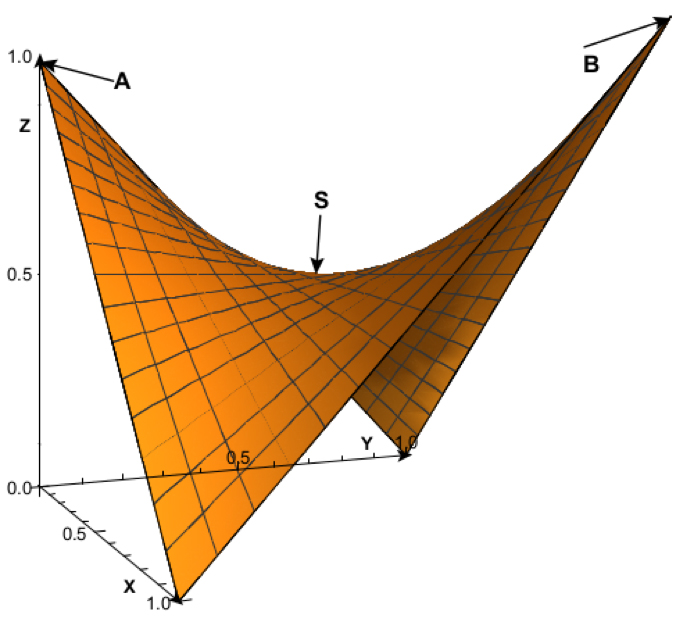
FIG. 2: Paraboloide hiperbòlic de la funció F’ de l’eficàcia biològica ponderada. L’eix X és la proporció de pressió selectiva individual p, l’eix I és la proporció d’eficàcia biològica individual e, i l’eix Z correspon al valor de F’. El punt S (0.5, 0.5, 0.5) és el punt de cadira o minimax. La paràbola que va del punt A (0,0,1) fins al punt B (1,1,1) i el vèrtex coincideix amb el punt de cadira S, representa els valors màxims d’e per a cada valor donat de p. És a dir, la paràbola ASB és la cresta de l’onada i representa les proporcions d’eficàcia que seran seleccionades positivament per a cada nivell de pressió selectiva.
D’aquest model destaquem que:
- Per a explicar l’emergència de l’altruisme evolutiu no cal el parentiu. Simplement amb la concurrència d’individus en un ambient en què la intensitat de la pressió de grup és superior a la individual s’han de seleccionar positivament trets que augmenten l’altruisme. En un ambient en què “p < 0,5”, els individus altruistes “e < 0,5” que treballen per al grup han de dotar-se de característiques cada vegada més altruistes, perquè els trets egoistes es converteixen en deleteris per al grup, i per tant faran que els individus tinguen una F’ menor.
- Encara que parega paradoxal, només serà beneficiosa per a l’individu la col·laboració dels seus companys si la pressió selectiva afavoreix l’egoisme “p > 0,5”.
- A llarg termini, quan predomine la pressió de grup “p < 0,5” i es mantinga aquest predomini en el temps, cada vegada seran seleccionats els més altruistes “e < 0,5”, i s’arribarà al punt màxim de la cresta de l’onada, el punt A (figura 1), on l’egoisme és nul i només es viu per al grup. Amb això s’arriba a la “individualització” del grup, a la simbiogènesi o especiació jeràrquica, o creació d’una nova espècie jeràrquicament superior. Al contrari, quan predomina i persisteix en el temps la pressió selectiva individual, es tendirà al punt B (figura 1) de màxim egoisme.
AUTOR
Javier Falgueras Cano
Premi “Accèssit” al XV Certamen Universitari Arquímedes pel treball “Altruisme” Realitzat per Javier Falgueras Cano de la Universitat de Burgos i tutoritzat per José Miguel Carretero Dia.
+INFO CERTAMEN ARQUÍMEDES



No comments yet.